 |
| Taças feitas com garrafas de Skol Beats. |
 |
| Vaso de flor feito com garrafa de Skol Beats. |
 |
| Enfeite de mesa feito com garrafa de Skol Beats. |
|
| Taças feitas com garrafas de Skol Beats. |
|
| Vaso de flor feito com garrafa de Skol Beats. |
|
| Enfeite de mesa feito com garrafa de Skol Beats. |
 |
| Primeiramente recolhemos e juntamos garrafas e latas de Skol Beats. |
 |
| Em seguida lavamos cada embalagem. |
 |
| Deixamos secar durante alguns dias. |

 |
| Depois veio a etapa de remoção da parte superior das latas e do corte nas garrafas. |



 |
A imagem mostra duas espécies de anêmonas, onde uma das espécies surgiu por poliploidia, ou seja, ocorreu a duplicação no número de cromossomos.
|
 |
Muitos estudiosos acreditam que diversas espécies de girassóis parecem ter surgidos desse modo, como mostram as imagens.
|
 |
Um exemplo é a mosca Rhagolettis pomonella, que seus ancestrais depositavam seus ovos, há 250 anos atrás, apenas nos frutos de um espinheiro da região do tipo Crataegus sp. Porém, mais tarde, com a introdução da maça doméstica, pelos europeus, na mesma região, parte dessa população passou a depositar seus ovos nessa maçã, resultando no acasalamento apenas entre as moscas ali presentes em cada fruto.
|
 |
Essa imagem mostra um esquema sobre a especiação parapátrica, onde envolve uma população de besouros. Como mostra a figura, é mais fácil que ocorra o cruzamento entre indivíduos que estão mais próximos do que os da mesma população que estão mais longes. Assim, mais pra frente a deriva genética e a seleção pode levar ao isolamento das mesmas.
|
 |
Por exemplo, vamos supor que uma espécie de larva fosse deixada em uma ilha (população parental), onde na mesma ilha já estivesse uma outra espécie de larva (população fundadora), com características diferentes. Mutações, recombinações, deriva genética e seleção proporcionariam mudanças genéticas e provocariam o isolamento das duas populações, ou seja, as moscas não conseguiriam mais cruzar e gerar novas larvas por serem tão diferentes.
|
 |
| A barreira geográfica pode ser um rio, uma montanha ou um cânion, por exemplo, como mostra o esquema. |


 |
| Uma população de pinguins pode ser entendida como uma população mendeliana, na qual os indivíduos se reproduzem sexuadamente e compartilham um conjunto gênico comum. |

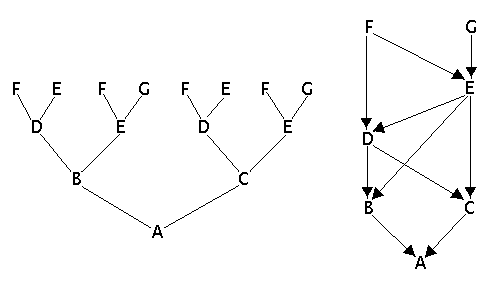
 |
| Esquema de mutação em um dor nucleotídios de um segmento de DNA. |
 |
| Exemplo de mimetismo. |
 |
| Coruja camuflada. |
 |
| Rã com coloração de aviso. |

 |
| Embriologia comparada de oito cordados. |


 |
| Representação de quatro estruturas homólogas. A correspondência entre os ossos dos animais está representada com a mesma cor. |
 |
| Representação de exemplo de divergência evolutiva. Um espécia ancestral do continente sul-americano originou as várias espécies que vivem nas ilhas do arquipélago de Galápagos. |
 |
| Fóssil de um dinossauro terrestre. |
 |
| Fóssil de um dinossauro marinho. |
 |
| Fóssil de um peixe. |
 |
| Gregor Mendel, o pai da genética. |
 |
| Esquema da mudança evolutiva: na concepção de Lamarck (A), ela ocorre no indivíduo; na teoria proposta por Darwin e Wallace (B), ela ocorre na população. |
 |
| Alfred Russel Wallace, naturalista britânico (1823-1913). |
